Механизм передачи возбуждения в вегетативных ганглиях. Вегетативная нервная система. Структурно – функциональные особенности.Синапсы, медиаторы и рецепторы ВНС. Синаптическая щель среди пост- и пресинаптических мембран
Нейроны
разделяются на три основных типа: афферентные, Эфферентные и промежуточные. Афферентные нейроны
(чувстви-гельные, или центростремительные) передают информацию от рецепторов в ц.н.с. Тела этих нейронов
расположены вне ц.н.с.- в спинномозговых узлах и в узлахчерепных нервов.Афферентный нейрон
имеют длинный отросток- дендрит, который контактирует на периферии с вопринимающем образованием- рецептором или сам образует рецептор, а также второй отросток - аксон, входящий через задние рога в спинной мозг. Эфферентные нейроны
(центробежные) связаны с передачей нисходящих влияний от вышележащих уровней нервной системы книжележащим или к рабочим органам. Например, нисходящие влиянияот пирамидных нейронов
коры больших полушарий или из других
моторных центров ц.н.с. следуют к нейронам спинного мозга (мотонейронам), от которых идут волокна к скелетным мышцам.В боковых рогах спинного мозга находятся клетки вегетативнойнервной системы, от которых идут пути к внутренним органам.Для эфферентных нейронов
характерны разветвлённая сеть коротких отростков -дендридов
и один длинный отросток-аксон.Промежуточные нейроны (интернейроны), или вставочные,- это, как правило, более мелкие клетки, осуществляющие связь между различными (в частности, афферентными и эфферентными) нейронами.
Они передают нервные влияния в горизонтальном (например, в пределах одного сегмента спинного мозга) и в вертикальном направлениях (например, из одного сегмента спинного мозга в другие выше или нижележащие сегменты). Благодаря многочисленным разветвлениям аксона промежуточные нейроны
могут одновременно возбуждать другие нейроны (таковы, например, звездчатые клетки коры).
Вегетативные ганглии подразделяются на паравертебральные, превертебральные и интрамуральные. Первые 2 вида характерны для симпатического отдела вегетативной нервной системы, а интрамуральные ганглии – для парасимпатической.
Вегетативный ганглий имеет соединительнотканную капсулу и строму. Ганглий содержит мультиполярные нейроны с эксцентрично расположенными округлыми ядрами и крупными ядрышками. Мультиполярные нейроны являются двигательными. Они окружены мантийной глией, но она расположена менее плотно, чем в спинальном ганглииПаравертебральные ганглии расположены по обе стороны от позвоночника, образуя симпатические цепочки (симпатический ствол).Превертебральные ганглии находятся спереди от аорты, образуя брюшное сплетение, которое состоит из 3-х типов узлов: чревный (солнечный), верхнебрыжеечный, нижнебрыжеечный. Их мультиполярные нейроны имеют множество дендритов, которые обильно ветвятся. Аксоны же образуют постганглионарные безмиелиновые волокна, идущие вглубь иннервируемых органов и образующие там аксосоматические синапсы.Большинство нейронов симпатических ганглиев содержат в мелких пузырьках катехоламины. Последние выявляются методом Хайка.Кроме мультиполярных нейронов в нервных ганглиях есть МИФ-клетки, то есть мелкие интенсивно флюорисцирующие клетки. Они имеют небольшой перикарион и короткие отростки. МИФ-клетки выделяют катехоламины, тормозят передачу импульса от преганглионарных нервных волокон на периферические нейроны ганглия.Нейроны интрамуральных ганглиев парасимпатического отдела являются холинергическими. Они выявляются методом Кюлле (реакция на ацетилхолин-эстеразу). Интрамуральные ганглии расположены в стенке органов и образуют сплетения, наиболее чётко выявленные в ЖКТ: подслизистое сплетение Мейснера, межмышечное сплетение Ауэрбаха и подсерозное сплетение Воробьёва.Нейроны интрамуральных ганглиев гетерогенны. Ниже представлена классификация данных нейронов по Догелю.Клетка Догеля I типа: длинноаксонный эффекторный нейрон. Перикарион уплощённый, много коротких дендритов с расширенными основанием, 1 длинный аксон. Аксон оканчивается на клетках-мишенях, на пример, гладких миоцитах.Клетка Догеля II типа: равноотростчатые афферентные нейроны (чувствительные). Перикарион овальной формы с гладкой поверхностью, отростки равнодлинные, аксоны образуют синапсы с клетками Догеля I типа, формируя местную рефлекторную дугу.Клетка Догеля III типа: ассоциативные нейроны, которые контактируют с соседними ганглиями. Тела овальной или полигональной формы, имеют 1 аксон и множество дендритов. Это клетки синтезируют различные нейромедиаторы.
Под вегетативной (от лат. vegetare – расти) деятельностью организма понимают работу внутренних органов, которая обеспечивает энергией и прочими необходимыми для существования компонентами все органы и ткани. В конце XIX века французский физиолог Клод Бернар (Bernard C.) пришёл к выводу, что "постоянство внутренней среды организма – залог его свободной и независимой жизни". Как отмечал он ещё в 1878 году, внутренняя среда организма подчиняется строгому контролю, удерживающему её параметры в определённых рамках. В 1929 году американский физиолог Уолтер Кэннон (Cannon W.) предложил обозначать относительное постоянство внутренней среды организма и некоторых физиологических функций термином гомеостаз (греч. homoios – равный и stasis – состояние). Есть два механизма сохранения гомеостаза: нервный и эндокринный. В этой главе будет рассмотрен первый из них.
11.1. Вегетативная нервная система
Вегетативная нервная система иннервирует гладкие мышцы внутренних органов, сердце и внешнесекреторные железы (пищеварительные, потовые и т.д.). Иногда эту часть нервной системы называют висцеральной (от лат. viscera – внутренности) и очень часто – автономной. Последнее определение подчёркивает важную особенность вегетативной регуляции: она происходит лишь рефлекторно, т.е. не осознаётся и не подчиняется произвольному контролю, тем самым принципиально отличаясь от соматической нервной системы, иннервирующей скелетные мышцы. В англоязычной литературе как правило используется термин автономная нервная система, в отечественной её чаще называют вегетативной.
В самом конце XIX века британский физиолог Джон Лэнгли (Langley J.) подразделил вегетативную нервную систему на три отдела: симпатический, парасимпатический и энтеральный. Эта классификация остаётся общепризнанной и в настоящее время (хотя в отечественной литературе энтеральный отдел, состоящий из нейронов межмышечного и подслизистого сплетений желудочно-кишечного тракта, довольно часто называют метасимпатическим). В этой главе рассматриваются первые два отдела вегетативной нервной системы. Кэннон обратил внимание на их разные функции: симпатический управляет реакциями борьбы или бегства (в английском рифмующемся варианте: fight or flight), а парасимпатический необходим для покоя и усвоения пищи (rest and digest). Швейцарский физиолог Вальтер Хесс (Hess W.) предложил называть симпатический отдел эрготропным, т.е. способствующим мобилизации энергии, интенсивной деятельности, а парасимпатический – трофотропным, т.е. регулирующим питание тканей, восстановительные процессы.
11.2. Периферический отдел вегетативной нервной системы
Прежде всего необходимо отметить, что периферический отдел вегетативной нервной системы является исключительно эфферентным, он служит только для проведения возбуждения к эффекторам. Если в соматической нервной системе для этого нужен всего лишь один нейрон (мотонейрон), то в вегетативной используются два нейрона, соединяющиеся через синапс в специальном вегетативном ганглии (Рис. 11.1).
Тела преганглионарных нейронов расположены в стволе мозга и спинном мозгу, а их аксоны направляются к ганглиям, где находятся тела постганглионарных нейронов. Рабочие органы иннервируются аксонами постганглионарных нейронов.

Симпатический и парасимпатический отделы вегетативной нервной системы отличаются прежде всего местом нахождения преганглионарных нейронов. Тела симпатических нейронов расположены в боковых рогах грудного и поясничного (два-три верхних сегмента) отделов. Преганглионарные нейроны парасимпатического отдела находятся, во-первых, в стволе мозга, откуда аксоны этих нейронов выходят в составе четырёх черепно-мозговых нервов: глазодвигательного (III), лицевого (VII), языкоглоточного (IX) и блуждающего (Х). Во-вторых, парасимпатические преганглионарные нейроны содержатся в крестцовом отделе спинного мозга (Рис. 11.2).
Симпатические ганглии принято подразделять на два типа: паравертебральные и превертебральные. Паравертебральные ганглии образуют т.н. симпатические стволы, состоящие из соединённых продольными волокнами узлов, которые располагаются по обе стороны от позвоночника на протяжении от основания черепа до крестца. В симпатическом стволе большинство аксонов преганглионарных нейронов передают возбуждение постганглионарным нейронам. Меньшая часть преганглионарных аксонов проходит через симпатический ствол к превертебральным ганглиям: шейным, звёздчатому, чревному, верхнему и нижнему брыжеечным – в этих непарных образованиях так же, как и в симпатическом стволе, находятся симпатические постганглионарные нейроны. Кроме того, часть симпатических преганглионарных волокон иннервирует мозговое вещество надпочечников. Аксоны преганглионарных нейронов тонкие и, несмотря на то, что многие из них покрыты миелиновой оболочкой, скорость проведения возбуждения по ним значительно меньше, чем по аксонам мотонейронов.
В ганглиях волокна преганглионарных аксонов ветвятся и образуют синапсы с дендритами многих постганглионарных нейронов (явление дивергенции), которые, как правило, мультиполярны и имеют в среднем около десятка дендритов. На один преганглионарный симпатический нейрон приходится в среднем около 100 постганглионарных нейронов. Вместе с тем, в симпатических ганглиях наблюдаются и конвергенция многих преганглионарных нейронов к одним и тем же постганглионарным. Благодаря этому происходит суммация возбуждения, а значит повышается надёжность передачи сигнала. Большинство симпатических ганглиев располагается достаточно далеко от иннервируемых органов и поэтому у постганглионарных нейронов довольно длинные аксоны, которые лишены миелинового покрытия.
В парасимпатическом отделе преганглионарные нейроны имеют длинные волокна, часть которых миелинизирована: они оканчиваются вблизи иннервируемых органов или в самих органах, где и находятся парасимпатические ганглии. Поэтому у постганглионарных нейронов аксоны оказываются короткими. Соотношение пре- и постганглионарных нейронов в парасимпатических ганглиях отличается от симпатических: оно составляет здесь лишь 1: 2. Большинство внутренних органов имеет как симпатическую, так и парасимпатическую иннервацию, важное исключение из этого правила составляют гладкие мышцы кровеносных сосудов, которые регулируются только симпатическим отделом. И лишь артерии половых органов имеют двойную иннервацию: и симпатическую, и парасимпатическую.
11.3. Тонус вегетативных нервов
Многие вегетативные нейроны обнаруживают фоновую спонтанную активность, т.е. способность самопроизвольно генерировать потенциалы действия в условиях покоя. Это означает, что иннервируемые ими органы при отсутствии какого-либо раздражения из внешней или внутренней среды всё равно получают возбуждение, обычно с частотой от 0,1 до 4 импульсов в секунду. Такая низкочастотная стимуляция по-видимому поддерживает постоянное небольшое сокращение (тонус) гладких мышц.
После перерезки или фармакологической блокады определённых вегетативных нервов иннервируемые органы лишаются их тонического влияния и такая утрата сразу же обнаруживается. Так, например, после односторонней перерезки симпатического нерва, контролирующего сосуды уха кролика, обнаруживается резкое расширение этих сосудов, а после перерезки или блокады блуждающих нервов у экспериментального животного учащаются сокращения сердца. Снятие блокады восстанавливает нормальную частоту сокращений сердца. После перерезки нервов частоту сокращений сердца и сосудистый тонус можно восстановить, если искусственно раздражать периферические отрезки электрическим током, подобрав его параметры так, чтобы они были близки к естественному ритму импульсации.
В результате различных влияний на вегетативные центры (что ещё предстоит рассмотреть в этой главе) их тонус может изменяться. Так, например, если по симпатическим нервам, контролирующим гладкие мышцы артерий, проходит 2 импульса в секунду, то ширина артерий типична для состояния покоя и тогда регистрируется нормальное артериальное давление. Если тонус симпатических нервов повысится и частота поступающих к артериям нервных импульсов увеличится, например до 4-6 в секунду, то гладкие мышцы сосудов будут сокращаться сильнее, просвет сосудов уменьшится, а артериальное давление возрастёт. И наоборот: при снижении симпатического тонуса частота поступающих к артериям импульсов становится меньше обычного, что приводит к расширению сосудов и понижению артериального давления.
Тонус вегетативных нервов имеет исключительно важное значение в регуляции деятельности внутренних органов. Он поддерживается благодаря поступлению к центрам афферентных сигналов, действию на них различных компонентов ликвора и крови, а также координирующему влиянию ряда структур головного мозга, в первую очередь – гипоталамуса.
11.4. Афферентное звено вегетативных рефлексов
Вегетативные реакции можно наблюдать при раздражении почти любой рецептивной области, но чаще всего они возникают в связи со сдвигами различных параметров внутренней среды и активацией интерорецепторов. Так, например, активация механорецепторов, находящихся в стенках полых внутренних органов (кровеносные сосуды, пищеварительный тракт, мочевой пузырь и т.д.) происходит при изменении в этих органах давления или объёма. Возбуждение хеморецепторов аорты и сонных артерий происходит вследствие повышения в артериальной крови напряжения углекислого газа или концентрации ионов водорода, а также при понижении напряжения кислорода. Осморецепторы активируются в зависимости от концентрации солей в крови или в ликворе, глюкорецепторы – в зависимости от концентрации глюкозы – любое изменение параметров внутренней среды вызывает раздражение соответствующих рецепторов и рефлекторную реакцию, направленную на сохранение гомеостаза. Во внутренних органах есть и болевые рецепторы, которые могут возбуждаться при сильном растяжении или сокращении стенок этих органов, при их кислородном голодании, при воспалении.
Интерорецепторы могут принадлежать одному из двух типов чувствительных нейронов. Во-первых, они могут быть чувствительными окончаниями нейронов спинальных ганглиев, и тогда возбуждение от рецепторов проводится, как обычно, в спинной мозг и затем, с помощью вставочных клеток, к соответствующим симпатическим и парасимпатическим нейронам. Переключения возбуждения с чувствительных на вставочные, а затем и эфферентные нейроны часто происходит в определённых сегментах спинного мозга. При сегментарной организации деятельность внутренних органов контролируют вегетативные нейроны, находящиеся в тех же самых сегментах спинного мозга, к которым поступает афферентная информация от этих органов.
Во-вторых, распространение сигналов от интерорецепторов может осуществляться по чувствительным волокнам, входящим в состав самих вегетативных нервов. Так, например, большая часть волокон, образующих блуждающий, языкоглоточный, чревный нервы, принадлежит не вегетативным, а чувствительным нейронам, тела которых находятся в соответствующих ганглиях.
11.5. Характер симпатического и парасимпатического влияния на деятельность внутренних органов
Большинство органов имеют двойную, т.е. симпатическую и парасимпатическую иннервацию. Тонус каждого из этих отделов вегетативной нервной системы может быть уравновешен влиянием другого отдела, но при определённых ситуациях обнаруживается повышенная активность, преобладание одного из них и тогда проявляется подлинный характер влияния этого отдела. Такое изолированное действие можно обнаружить и в экспериментах с перерезкой или фармакологической блокадой симпатических или парасимпатических нервов. После такого вмешательства деятельность рабочих органов изменяется под влиянием сохранившего с ним связь отдела вегетативной нервной системы. Другой способ экспериментального изучения состоит в поочерёдном раздражении симпатических и парасимпатических нервов специально подобранными параметрами электрического тока – этим моделируется повышение симпатического или парасимпатического тонуса.
Влияние двух отделов вегетативной нервной системы на управляемые органы чаще всего противоположно по направленности сдвигов, что даже даёт повод говорить об антагонистическом характере отношений симпатического и парасимпатического отделов. Так, например, при активации симпатических нервов, управляющих работой сердца, происходит увеличение частоты и силы его сокращений, растёт возбудимость клеток проводящей системы сердца, а при повышении тонуса блуждающих нервов регистрируются противоположные сдвиги: частота и сила сердечных сокращений уменьшаются, возбудимость элементов проводящей системы снижается. Другие примеры противоположного влияния симпатических и парасимпатических нервов можно видеть в таблице11.1

Несмотря на то, что влияние симпатического и парасимпатического отделов на многие органы оказывается противоположным, они действуют как синергисты, т.е. содружественно. При повышении тонуса одного из этих отделов синхронно снижается тонус другого: это означает, что физиологические сдвиги любой направленности обусловлены согласованными изменениями активности обоих отделов.
11.6. Передача возбуждения в синапсах вегетативной нервной системы
В вегетативных ганглиях и симпатического, и парасимпатического отделов медиатором является одно и то же вещество – ацетилхолин (Рис. 11.3). Этот же медиатор служит химическим посредником для передачи возбуждения от парасимпатических постганглионарных нейронов к рабочим органам. Основным медиатором симпатических постганглионарных нейронов является норадреналин.

Хотя в вегетативных ганглиях и в передаче возбуждения от парасимпатических постганглионарных нейронов на рабочие органы используется один и тот же медиатор, взаимодействующие с ним холинорецепторы не одинаковы. В вегетативных ганглиях с медиатором взаимодействуют никотин-чувствительные или Н-холинорецепторы. Если в эксперименте смочить клетки вегетативных ганглиев 0,5% раствором никотина, то они перестают проводить возбуждение. К такому же результату приводит введение раствора никотина в кровь экспериментальных животных и создание, тем самым, высокой концентрации этого вещества. В малой же концентрации никотин действует подобно ацетилхолину, т.е. возбуждает этот тип холинорецепторов. Такие рецепторы связаны с ионотропными каналами и при их возбуждении открываются натриевые каналы постсинаптической мембраны.
Холинорецепторы, находящиеся в рабочих органах и взаимодействующие с ацетилхолином постганглионарных нейронов, принадлежат к другому типу: они не реагируют на никотин, зато их можно возбудить малым количеством другого алкалоида – мускарина или блокировать высокой концентрацией этого же вещества. Мускарин-чувствительные или М-холинорецепторы обеспечивают метаботропное управление, в котором участвуют вторичные посредники, а вызываемые действием медиатора реакции развиваются медленнее и сохраняются дольше, чем при ионотропном управлении.
Медиатор симпатических постганглионарных нейронов норадреналин может связываться метаботропными адренорецепторами двух типов: a- или b, соотношение которых в разных органах не одинаково, что и определяет различные физиологические реакции на действие норадреналина. Например, в гладких мышцах бронхов преобладают b-адренорецепторы: действие медиатора на них сопровождается расслаблением мышц, что ведёт к расширению бронхов. В гладких мышцах артерий внутренних органов и кожи больше a-адренорецепторов и здесь мышцы под действием норадреналина сокращаются, что ведёт к сужению этих сосудов. Секрецию потовых желёз контролируют особые, холинэргические симпатические нейроны, медиатором которых является ацетилхолин. Есть сведения и о том, что артерии скелетных мышц тоже иннервируют симпатические холинэргические нейроны. Согласно другой точке зрения артерии скелетных мышц управляются адренэргическими нейронами, причём норадреналин действует на них через a-адренорецепторы. А тот факт, что при мышечной работе, всегда сопровождающейся повышением симпатической активности, артерии скелетных мышц расширяются, объясняют действием гормона мозгового вещества надпочечников адреналина на b-адренорецепторы
При симпатической активации адреналин в больших количествах выделяется из мозгового вещества надпочечников (следует обратить внимание на иннервацию мозгового вещества надпочечников симпатическими преганглионарными нейронами), и тоже взаимодействует с адренорецепторами. Это усиливает симпатическую реакцию, поскольку кровь приносит адреналин и к тем клеткам, вблизи которых нет окончаний симпатических нейронов. Норадреналин и адреналин стимулируют расщепление гликогена в печени и липидов в жировой ткани, действуя там на b-адренорецепторы. В сердечной мышце b-рецепторы намного чувствительнее к норадреналину, чем к адреналину, тогда как в сосудах и бронхах их легче активирует адреналин. Эти различия послужили основанием для разделения b-рецепторов на два типа: b1 (в сердце) и b2 (в других органах).
Медиаторы вегетативной нервной системы могут действовать не только на постсинаптическую, но и на пресинаптическую мембрану, где тоже имеются соответствующие рецепторы. Пресинаптические рецепторы используются для регуляции количества выделяемого медиатора. Например, при повышенной концентрации норадреналина в синаптической щели он действует на пресинаптические a-рецепторы, что приводит к уменьшению его дальнейшего выделения из пресинаптического окончания (отрицательная обратная связь). Если же концентрация медиатора в синаптической щели становится низкой, с ним взаимодействуют преимущественно b-рецепторы пресинаптической мембраны, а это ведёт к повышению выделения норадреналина (положительная обратная связь).
По такому же принципу, т.е. с участием пресинаптических рецепторов, осуществляется регуляция выделения ацетилхолина. Если окончания симпатических и парасимпатических постганглионарных нейронов оказываются поблизости друг от друга, то возможно реципрокное влияние их медиаторов. Например, пресинаптические окончания холинэргических нейронов содержат a-адренорецепторы и, если на них подействует норадреналин, то выделение ацетилхолина уменьшится. Таким же образом ацетилхолин может уменьшать выделение норадреналина, если присоединится к М-холинорецепторам адренэргического нейрона. Таким образом, симпатический и парасимпатический отделы конкурируют даже на уровне постганглионарных нейронов.
Очень многие лекарственные препараты действуют на передачу возбуждения в вегетативных ганглиях (ганглиоблокаторы, a-адреноблокаторы, b-блокаторы и т.д.) и поэтому широко применяются в медицинской практике для коррекции различного рода нарушений вегетативной регуляции.
11.7. Центры вегетативной регуляции спинного мозга и ствола
Многие преганглионарные и постганглионарные нейроны способны активироваться независимо друг от друга. Например, одни симпатические нейроны управляют потоотделением, а другие – кожным кровотоком, секрецию слюнных желёз повышают одни парасимпатические нейроны, а секрецию железистых клеток желудка – другие. Существуют такие методы обнаружения активности постганглионарных нейронов, которые позволяют отличить сосудосуживающие нейроны кожи от холинэргических нейронов, управляющих сосудами скелетных мышц или от нейронов, действующих на волосковые мышцы кожи.
Топографически организованный вход афферентных волокон от разных рецептивных областей к определённым сегментам спинного мозга или разным областям ствола возбуждает вставочные нейроны, а они передают возбуждение преганглионарным вегетативным нейронам, замыкая таким образом рефлекторную дугу. Наряду с этим для вегетативной нервной системы характерна интегративная деятельность, которая особенно выражена в симпатическом отделе. При определённых обстоятельствах, например, при переживании эмоций может повышаться активность всего симпатического отдела, а соответственно этому снижается активность парасимпатических нейронов. Кроме того, активность вегетативных нейронов согласуется с деятельностью мотонейронов, от которых зависит работа скелетных мышц, но их снабжение необходимыми для работы глюкозой и кислородом осуществляется под контролем вегетативной нервной системы. Участие вегетативных нейронов в интегративной деятельности обеспечивают вегетативные центры спинного мозга и ствола.
В грудном и поясничном отделах спинного мозга находятся тела симпатических преганглионарных нейронов, которые образуют промежуточно-боковое, вставочное и небольшое центрально-вегетативное ядра. Симпатические нейроны, контролирующие потовые железы, сосуды кожи и скелетных мышц располагаются латерально по отношению к нейронам, регулирующим деятельность внутренних органов. По такому же принципу расположены в крестцовом отделе спинного мозга парасимпатические нейроны: латерально – иннервирующие мочевой пузырь, медиально – толстый кишечник. После отделения спинного мозга от головного вегетативные нейроны способны ритмически разряжаться: например, симпатические нейроны двенадцати сегментов спинного мозга, объединённые внутриспинальными проводящими путями, могут, в определённой степени, рефлекторно регулировать тонус кровеносных сосудов. Однако у спинальных животных число разряжающихся симпатических нейронов и частота разрядов оказываются меньше, чем у интактных. Это значит, что контролирующие тонус сосудов нейроны спинного мозга стимулируются не только афферентным входом, но и центрами головного мозга.
В стволе мозга находятся сосудодвигательный и дыхательный центры, которые ритмически активируют симпатические ядра спинного мозга. К стволу непрерывно поступает афферентная информация от баро- и хеморецепторов и в соответствии с её характером вегетативные центры определяют изменения тонуса не только симпатических, но и парасимпатических нервов, контролирующих, например, работу сердца. Это рефлекторная регуляция, в которую вовлекаются и мотонейроны дыхательных мышц – они ритмически активируются дыхательным центром.
В ретикулярной формации мозгового ствола, где расположены вегетативные центры, используется несколько медиаторных систем, осуществляющих контроль важнейших гомеостатических показателей и находящихся в сложных отношениях между собой. Здесь одни группы нейронов могут стимулировать деятельность других, тормозить активность третьих и одновременно испытывать влияние и тех, и других на себе. Наряду с центрами регуляции кровообращения и дыхания здесь находятся нейроны, координирующие многие пищеварительные рефлексы: слюноотделение и глотание, выделение желудочного сока, моторику желудка; отдельно можно упомянуть защитный рвотный рефлекс. Разные центры постоянно координируют свою деятельность друг с другом: например, при глотании рефлекторно закрывается вход в дыхательные пути и, благодаря этому, предупреждается вдох. Активность стволовых центров подчиняет себе деятельность вегетативных нейронов спинного мозга.
11. 8. Роль гипоталамуса в регуляции вегетативных функций
На гипоталамус приходится менее 1% объёма мозга, однако он играет решающую роль в регуляции вегетативных функций. Это объясняется несколькими обстоятельствами. Во-первых, гипоталамус оперативно получает информацию от интерорецепторов, сигналы от которых поступают к нему через ствол мозга. Во-вторых, сюда приходит информация от поверхности тела и от ряда специализированных сенсорных систем (зрительная, обонятельная, слуховая). В-третьих, некоторые нейроны гипоталамуса имеют собственные осмо-, термо- и глюкорецепторы (такие рецепторы называются центральными). Они могут реагировать на сдвиги осмотического давления, температуры и уровня глюкозы в ликворе и крови. В связи с этим следует напомнить, что в гипоталамусе меньше, по сравнению с остальным мозгом, проявляются свойства гематоэнцефалического барьера. В-четвёртых, гипоталамус имеет двусторонние связи с лимбической системой мозга, ретикулярной формацией и корой больших полушарий, что позволяет ему координировать вегетативные функции с определённым поведением, например, с переживанием эмоций. В-пятых, гипоталамус образует проекции на вегетативные центры ствола и спинного мозга, что позволяет ему осуществлять прямой контроль деятельности этих центров. В-шестых, гипоталамус контролирует важнейшие механизмы эндокринной регуляции (См. главу 12).
Важнейшие для вегетативной регуляции переключения осуществляют нейроны ядер гипоталамуса (Рис. 11.4), в разных классификациях их насчитывают от 16 до 48. В 40-х годах ХХ века Вальтер Хесс (Hess W.) через введённые с помощью стереотаксической техники электроды последовательно раздражал разные области гипоталамуса у экспериментальных животных и обнаружил при этом разные комбинации вегетативных и поведенческих реакций.

При стимуляции задней области гипоталамуса и прилегающего к водопроводу серого вещества у подопытных животных повышалось кровяное давление, увеличивалась частота сокращений сердца, учащалось и углублялось дыхание, расширялись зрачки, а также поднималась шерсть, изгибалась горбом спина и оскаливались зубы, т.е. вегетативные сдвиги говорили об активации симпатического отдела, а поведение было аффективно-оборонительным. Раздражение ростральных отделов гипоталамуса и преоптической области вызывало у тех же животных пищевое поведение: они начинали есть, даже если были досыта накормлены, при этом увеличивалось выделение слюны и усиливалась моторика желудка и кишечника, а частота сердечных сокращений и дыхания уменьшалась, становился меньшим и мышечный кровоток, что вполне характерно для повышения парасимпатического тонуса. Одну область гипоталамуса с лёгкой руки Хесса стали называть эрготропной, а другую – трофотропной; их отделяет друг от друга каких-нибудь 2-3 мм.
Из этих и многих других исследований постепенно сложилось представление о том, что активация разных областей гипоталамуса запускает уже предуготованный комплекс поведенческих и вегетативных реакций, а значит роль гипоталамуса состоит в том, чтобы оценить поступающую к нему из разных источников информацию и на её основе выбрать тот или иной вариант, объединяющий поведение с определённой активностью обоих отделов вегетативной нервной системы. Само же поведение можно в этой ситуации рассматривать как деятельность, направленную на предупреждение возможных сдвигов внутренней среды. Следует обратить внимание, что не только уже произошедшие отклонения гомеостаза, но и любое потенциально угрожающее гомеостазу событие может активировать необходимую деятельность гипоталамуса. Так, например, при внезапной угрозе вегетативные сдвиги у человека (увеличение частоты сокращений сердца, повышение кровяного давления и т.п.) происходят быстрее, чем он обратится в бегство, .т.е. такие сдвиги уже учитывают характер последующей мышечной активности.
Непосредственный контроль тонуса вегетативных центров, а значит и выходной активности вегетативной нервной системы, гипоталамус осуществляет с помощью эфферентных связей с тремя важнейшими областями (Рис. 11.5):

1). Ядро солитарного тракта в верхнем отделе продолговатого мозга, которое является главным адресатом сенсорной информации от внутренних органов. Оно взаимодействует с ядром блуждающего нерва и других парасимпатических нейронов и участвует в контроле температуры, кровообращения и дыхания. 2). Ростральная вентральная область продолговатого мозга, имеющая решающее значение в повышении общей выходной активности симпатического отдела. Эта активность проявляется в повышении кровяного давления, увеличении частоты сокращений сердца, секреции потовых желёз, расширении зрачков и сокращении мышц, поднимающих волосы. 3). Вегетативные нейроны спинного мозга, на которые гипоталамус может оказывать прямое влияние.
11.9. Вегетативные механизмы регуляции кровообращения
В замкнутой сети кровеносных сосудов и сердца (Рис. 11.6) постоянно движется кровь, объём которой составляет в среднем 69 мл/кг массы тела у взрослых мужчин и 65 мл/кг массы тела у женщин (т.е. при массе тела 70 кг он составит соответственно 4830 мл и 4550 мл). В состоянии покоя от 1/3 до 1/2 этого объёма не циркулирует по сосудам, а находится в кровяных депо: капиллярах и венах брюшной полости, печени, селезёнки, лёгких, подкожных сосудов.

При физической работе, эмоциональных реакциях, стрессе эта кровь переходит из депо в общий кровоток. Движение крови обеспечивают ритмические сокращения желудочков сердца, каждое из которых изгоняет в аорту (левый желудочек) и лёгочную артерию (правый желудочек) приблизительно по 70 мл крови, а при тяжёлой физической нагрузке у хорошо тренированных людей этот показатель (его называют систолическим или ударным объёмом) может увеличиваться до 180 мл. Сердце взрослого человека сокращается в покое приблизительно 75 раз в минуту, а значит за это время через него должно пройти свыше 5 литров крови (75´70 = 5250 мл) – этот показатель называется минутным объёмом кровообращения. При каждом сокращении левого желудочка давление в аорте, а затем в артериях повышается до 100-140 мм рт. ст. (систолическое давление), а к началу следующего сокращения опускается до 60-90 мм (диастолическое давление). В лёгочной артерии эти показатели меньше: систолическое – 15-30 мм, диастолическое – 2-7 мм – это связано с тем, что т.н. малый круг кровообращения, начинающийся от правого желудочка и доставляющий кровь к лёгким, короче большого, а потому оказывает меньшее сопротивление току крови и не требует высокого давления. Таким образом, главными показателями функции кровообращения оказываются частота и сила сердечных сокращений (от неё зависит систолический объём), систолическое и диастолическое давление, которые определяются объёмом жидкости в замкнутой системе кровобращения, минутным объёмом кровотока и сопротивлением сосудов этому кровотоку. Сопротивление сосудов изменяется в связи с сокращениями их гладких мышц: чем уже становится просвет сосуда, тем большее сопротивление кровотоку он оказывает.
Постоянство объёма жидкости в организме регулируют гормоны (См. главу 12), но какая часть крови будет находиться в депо, а какая циркулировать по сосудам, какое сопротивление окажут сосуды кровотоку – зависит от управления сосудами симпатическим отделом. Работу сердца, а значит и величину артериального давления, в первую очередь систолического, контролируют и симпатические, и блуждающие нервы (хотя эндокринные механизмы и местная саморегуляция здесь тоже играют важную роль). Механизм слежения за изменениями важнейших параметров системы кровообращения довольно прост, он сводится к непрерывной регистрации барорецепторами степени растяжения дуги аорты и места разделения общих сонных артерий на наружные и внутренние (эта область называется каротидным синусом). Этого достаточно, поскольку растяжение указанных сосудов отражает и работу сердца, и сопротивление сосудов, и объём крови.
Чем сильнее растягиваются аорта и сонные артерии, тем с большей частотой распространяются от бароцепторов нервные импульсы по чувствительным волокнам языкоглоточного и блуждающего нервов к соответствующим ядрам продолговатого мозга. Это приводит к двум последствиям: повышению влияния блуждающего нерва на сердце и понижению симпатического влияния на сердце и сосуды. В результате уменьшается работа сердца (снижается минутный объём) и понижается тонус сосудов, оказывающих сопротивление кровотоку, а это приводит к уменьшению растяжения аорты и сонных артерий и соответственному уменьшению импульсации от барорецепторов. Если она станет понижаться, то произойдёт повышение симпатической активности и снизится тонус блуждающих нервов, а в результате опять восстановится надлежащее значение важнейших параметров кровообращения.
Непрерывное движение крови необходимо, в первую очередь, для того, чтобы доставлять работающим клеткам кислород от лёгких, а образующийся в клетках углекислый газ уносить к лёгким, где он выделяется из организма. Содержание этих газов в артериальной крови поддерживается на неизменном уровне, который отражают значения их парциального давления (от лат. pars – часть, т.е. частичного от целого атмосферного): кислорода – 100 мм рт. ст., углекислого газа – около 40 мм рт. ст. Если ткани станут работать интенсивнее, то они начнут забирать из крови больше кислорода и отдавать в неё больше углекислого газа, что приведёт соответственно к понижению содержания кислорода и повышению – углекислого газа в артериальной крови. Эти сдвиги улавливают хеморецепторы, расположенные в тех же сосудистых областях, что и барорецепторы, т.е. в аорте и развилках сонных артерий, питающих мозг. Поступление более частых сигналов от хеморецепторов в продолговатый мозг приведёт к активации симпатического отдела и уменьшению тонуса блуждающих нервов: в результате усилится работа сердца, повысится тонус сосудов и под большим давлением кровь станет быстрее циркулировать между лёгкими и тканями. Одновременно с этим увеличенная по частоте импульсация от хеморецепторов сосудов приведёт к учащению и углублению дыхания и быстро циркулирующая кровь станет быстрее насыщаться кислородом и освобождаться от излишков углекислого газа: в итоге показатели газового состава крови нормализуются.
Таким образом, барорецепторы и хеморецепторы аорты и сонных артерий незамедлительно реагируют на сдвиги гемодинамических параметров (проявляющиеся увеличением или уменьшением растяжения стенок этих сосудов), а также на изменения насыщения крови кислородом и углекислым газом. Вегетативные центры, получившие от них информацию, так изменяют тонус симпатического и парасимпатического отделов, что оказываемое ими на рабочие органы влияние приводит к нормализации отклонившихся от гомеостатических констант параметров.
Конечно, это лишь часть сложной системы регуляции кровообращения, в которой наряду с нервными существуют ещё гуморальные и местные механизмы регуляции. Например, любой особенно интенсивно работающий орган потребляет больше кислорода и образует больше недоокисленных продуктов обмена, которые способны сами расширять сосуды, снабжающие орган кровью. В результате он начинает забирать из общего потока крови больше, чем забирал прежде, а потому в центральных сосудах из-за уменьшающегося объёма крови понижается давление и возникает необходимость регулировать этот сдвиг уже с помощью нервных и гуморальных механизмов.
При физической работе система кровообращения должна прилаживаться и к мышечным сокращениям, и к повышенному потреблению кислорода, и к накоплению продуктов обмена, и к изменяющейся активности других органов. При различных поведенческих реакциях, при переживании эмоций в организме происходят сложные изменения, отражающиеся на постоянстве внутренней среды: в таких случаях весь комплекс таких изменений, активирующих разные области мозга, непременно отражается на активности нейронов гипоталамуса, а он уже координирует механизмы вегетативной регуляции с мышечной работой, эмоциональным состоянием или поведенческими реакциями.
11.10. Основные звенья регуляции дыхания
При спокойном дыхании в лёгкие во время вдоха входит около 300-500 куб. см воздуха и такой же объём воздуха при выдохе уходит в атмосферу – это т.н. дыхательный объём. После спокойного вдоха можно дополнительно вдохнуть 1,5-2 литра воздуха – это резервный объём вдоха, а после обычного выдоха можно изгнать из лёгких ещё 1-1,5 литра воздуха – это резервный объём выдоха. Сумма дыхательного и резервных объёмов составляет т.н. жизненную ёмкость лёгких, которую обычно определяют с помощью спирометра. Взрослые люди дышат в среднем 14-16 раз в минуту, вентилируя за это время через лёгкие 5-8 литров воздуха – это минутный объём дыхания. При увеличении глубины дыхания за счёт резервных объёмов и одновременном повышении частоты дыхательных движений можно в несколько раз увеличить минутную вентиляцию лёгких (в среднем до 90 литров в минуту, а тренированные люди способны удвоить и этот показатель).
Воздух поступает в альвеолы лёгких – воздушные ячейки, густо оплетённые сетью кровеносных капилляров, несущих венозную кровь: она мало насыщена кислородом и избыточно – углекислым газом (Рис. 11.7).

Очень тонкие стенки альвеол и капилляров не препятствуют газообмену: по градиенту парциальных давлений кислород из альвеолярного воздуха переходит в венозную кровь, а углекислый газ диффундирует в альвеолы. В результате от альвеол течёт артериальная кровь с парциальным давлением в ней кислорода около 100 мм рт. ст., а углекислого газа – не более 40 мм рт. ст.. Вентиляция лёгких всё время обновляет состав альвеолярного воздуха, а непрерывный кровоток и диффузия газов через лёгочную мембрану позволяют постоянно превращать венозную кровь в артериальную.
Вдох происходит благодаря сокращениям дыхательных мышц: наружных межрёберных и диафрагмы, которые управляются двигательными нейронами шейного (диафрагма) и грудного отдела спинного мозга (межрёберные мышцы). Эти нейроны активируются нисходящими из дыхательного центра ствола мозга путями. Дыхательный центр образуют несколько групп нейронов продолговатого мозга и моста, одна из них (дорсальная инспираторная группа) самопроизвольно активируется в условиях покоя 14-16 раз в минуту, а это возбуждение проводится к двигательным нейронам дыхательных мышц. В самих лёгких, в покрывающей их плевре и в воздухоносных путях есть чувствительные нервные окончания, которые возбуждаются при растяжении лёгких и движении воздуха по дыхательным путям во время вдоха. Сигналы от этих рецепторов поступают в дыхательный центр, который на их основе регулирует продолжительность и глубину вдоха.
При нехватке кислорода в воздухе (например, в разряжённом воздухе горных вершин) и при физической работе насыщение крови кислородом уменьшается. При физической работе одновременно с этим растёт содержание углекислого газа в артериальной крови, поскольку лёгкие, работая в обычном режиме, не успевают очищать от него кровь до необходимой кондиции. На сдвиг газового состава артериальной крови реагируют хеморецепторы аорты и сонных артерий, сигналы от которых поступают к дыхательному центру. Это приводит к изменению характера дыхания: вдох происходит чаще и делается глубже за счёт резервных объёмов, выдох, обычно пассивный, становится при таких обстоятельствах форсированным (активируется вентральная группа нейронов дыхательного центра и начинают действовать внутренние межрёберные мышцы). В результате этого увеличивается минутный объём дыхания и большая вентиляция лёгких при одновременно увеличенном потоке крови через них позволяет восстановить газовый состав крови до гомеостатического стандарта. Сразу после интенсивной физической работы у человека сохраняется одышка и учащённый пульс, которые прекращаются, когда кислородный долг будет погашен.
Ритм активности нейронов дыхательного центра приспосабливается и к ритмической деятельности дыхательных, и других скелетных мышц, от проприоцепторов которых он непрерывно получает информацию. Координацию дыхательной ритмики с другими гомеостатическими механизмами осуществляет гипоталамус, он же, взаимодействуя с лимбической системой и корой, меняет модель дыхания при эмоциональных реакциях. Кора больших полушарий может оказывать прямое влияние на функцию дыхания, приспосабливая его к разговору или пению. Только непосредственное влияние коры позволяет произвольно изменять характер дыхания, намеренно задерживать его, урежать или учащать, но всё это возможно лишь в ограниченных пределах. Так, например, произвольная задержка дыхания у большинства людей не превышает минуты, после чего оно непроизвольно возобновляется из-за чрезмерного накопления углекислого газа в крови и одновременного уменьшения в ней кислорода.
Резюме
Постоянство внутренней среды организма является гарантом его свободной деятельности. Быстрое восстановление смещённых гомеостатических констант осуществляет вегетативная нервная система. Она способна также предупреждать возможные сдвиги гомеостаза, связанные с изменениями внешней среды. Два отдела вегетативной нервной системы одновременно контролируют деятельность большинства внутренних органов, оказывая на них противоположное влияние. Повышение тонуса симпатических центров проявляется эрготропными реакциями, а повышение парасимпатического тонуса – трофотропными. Активность вегетативных центров координирует гипоталамус, он согласует их деятельность с работой мышц, эмоциональными реакциями и поведением. Гипоталамус взаимодействует с лимбической системой мозга, ретикулярной формацией и корой больших полушарий. Вегетативные механизмы регуляции играют главную роль в осуществлении жизненно важных функций кровообращения и дыхания.
Вопросы для самоконтроля
165. В каком отделе спинного мозга находятся тела парасимпатических нейронов?
А. Шейный; Б. Грудной; В. Верхние сегменты поясничного отдела; Г. Нижние сегменты поясничного отдела; Д. Крестцовый.
166. Какие черепно-мозговые нервы не содержат волокон парасимпатических нейронов?
А. Тройничные; Б. Глазодвигательные; В. Лицевые; Г. Блуждающие; Д. Языкоглоточные.
167. Какие ганглии симпатического отдела следует отнести к паравертебральным?
А. Симпатический ствол; Б. Шейный; В. Звёздчатый; Г. Чревный; В. Нижний брыжеечный.
168. Какой из указанных ниже эффекторов получает в основном лишь симпатическую иннервацию?
А. Бронхи; Б. Желудок; В. Кишечник; Г. Кровеносные сосуды; Д. Мочевой пузырь.
169. Что из перечисленного отражает повышение тонуса парасимпатического отдела?
А. Расширение зрачков; Б. Расширение бронхов; В. Повышение частоты сокращений сердца; Г. Повышение секреции пищеварительных желёз; Д. Повышение секреции потовых желёз.
170. Что из указанного характерно для повышения тонуса симпатического отдела?
А. Повышение секреции бронхиальных желёз; Б. Усиление моторики желудка; В. Повышение секреции слёзных желёз; Г. Сокращение мускулатуры мочевого пузыря; Д. Увеличенное расщепление углеводов в клетках.
171. Деятельность какой эндокринной железы контролируется симпатическими преганглионарными нейронами?
А. Кора надпочечников; Б. Мозговое вещество надпочечников; В. Поджелудочная железа; Г. Щитовидная железа; Д. Околощитовидные железы.
172. С помощью какого нейромедиатора происходит передача возбуждения в симпатических вегетативных ганглиях?
А. Адреналин; Б. Норадреналин; В. Ацетилхолин; Г. Дофамин; Д. Серотонин.
173. С помощью какого медиатора парасимпатические постганглионарные нейроны обычно действуют на эффекторы?
А. Ацетилхолин; Б. Адреналин; В. Норадреналин; Г. Серотонин; Д. Вещество Р.
174. Что из указанного характеризует Н-холинорецепторы?
А. Принадлежат постсинаптической мембране рабочих органов, регулируемых парасимпатическим отделом; Б. Ионотропные; В. Активируются мускарином; Г. Относятся только к парасимпатическому отделу; Д. Находятся только на пресинаптической мембране.
175. Какие рецепторы должны связаться с медиатором, чтобы в эффекторной клетке началось повышенное расщепление углеводов?
А. a-адренорецепторы; Б. b-адренорецепторы; В. Н-холинорецепторы; Г. М-холинорецепторы; Д. Ионотропные рецепторы.
176. Какая структура мозга осуществляет координацию вегетативных функций и поведения?
А. Спинной мозг; Б. Продолговатый мозг; В. Средний мозг; Г. Гипоталамус; Д. Кора больших полушарий.
177. Какой гомеостатический сдвиг окажет непосредственное действие на центральные рецепторы гипоталамуса?
А. Повышение артериального давления; Б. Повышение температуры крови; В. Увеличение объёма крови; Г. Повышение парциального давления кислорода в артериальной крови; Д. Снижение кровяного давления.
178. Чему равна величина минутного объёма кровообращения, если ударный объём равен 65 мл, а частота сокращений сердца – 78 в одну минуту?
А. 4820 мл; Б. 4960 мл; В. 5070 мл; Г. 5140 мл; Д. 5360 мл.
179. Где расположены барорецепторы, поставляющие информацию вегетативным центрам продолговатого мозга, осуществляющим регуляцию работы сердца и артериального давления?
А. Сердце; Б. Аорта и сонные артерии; В. Крупные вены; Г. Мелкие артерии; Д. Гипоталамус.
180. В положении лёжа у человека рефлекторно уменьшается частота сокращений сердца и артериальное давление. Активация каких рецепторов вызывает эти изменения?
А. Интрафузальные рецепторы мышц; Б. Сухожильные рецепторы Гольджи; В.Вестибулярные рецепторы; Г. Механорецепторы дуги аорты и сонных артерий; Д. Внутрисердечные механорецепторы.
181. Какое событие скорее всего произойдёт вследствие повышения напряжения углекислого газа в крови?
А. Уменьшение частоты дыхания; Б. Уменьшение глубины дыхания; В. Уменьшение частоты сокращений сердца; Г. Уменьшение силы сокращений сердца; Д. Повышение артериального давления.
182. Чему равна жизненная ёмкость лёгких, если дыхательный объём составляет 400 мл, резервный объём вдоха – 1500 мл, а резервный объём выдоха – 2 л?
А. 1900 мл; Б. 2400 мл; В. 3,5 л; Г. 3900 мл; Д. По имеющимся данным жизненную ёмкость лёгких определить невозможно.
183. Что может произойти вследствие непродолжительной произвольной гипервентиляции лёгких (частого и глубокого дыхания)?
А. Повышение тонуса блуждающих нервов; Б. Повышение тонуса симпатических нервов; В. Повышение импульсация от сосудистых хеморецепторов; Г. Повышение импульсация от барорецепторов сосудов; Д. Повышение систолического давления.
184. Что понимают под тонусом вегетативных нервов?
А. Их способность возбуждаться при действии раздражителя; Б. Способность проводить возбуждение; В. Наличие спонтанной фоновой активности; Г. Повышение частоты проводимых сигналов; Д. Любое изменение частоты передаваемых сигналов.
text_fields
text_fields
arrow_upward
Преганглионарные синапсы образуются нервными отростками вставочных нейронов вегетативных центров на нервных клетках вегетативных ганглиев. Число нейронов в вегетативном ганглии очень велико и в 2-30 раз превышает количество входящих в ганглий преганглионарных проводников. Поэтому каждое преганглионарное волокно ветвится и образует синапсы на нескольких нейронах ганглия. В то же время, на каждом нейроне ганглия имеются многочисленные синапсы разных преганглионарных волокон. Эти особенности обеспечивают нейронам ганглиев высокую способность к пространственной и временной суммации возбуждений.
Преганглионарные синапсы отличают три особенности:
1) значительная синаптическая задержка проведения, примерно в 5 раз продолжительнее, чем в центральных синапсах,
2) существенно большая длительность ВПСП,
3) наличие выраженной и продолжительной следовой гиперполяризации нейронов ганглия. Благодаря этим особенностям преганглионарные синапсы обладают невысокой лабильностью и обеспечивают трансформацию ритма возбуждений с частотой импульсации в постганглионарном волокне не более 15/с.
Медиатором во всех преганглионарных синапсах и симпатического, и парасимпатического отделов вегетативной нервной системы является ацетилхолин. Химические клеточные рецепторы постсинаптической мембраны, связывающие ацетилхолин, называют холинорецепторами и относят в преганглионарных синапсах к никотиночувствительным, так как они активируются никотином (Н-холинорецепторы). Специфическими блокаторами этих рецепторов являются кураре и курареподобные вещества (бензогексоний, дитилин и др.), входящие В группу ганглиоблокаторов. Кроме основных, участвующих в передаче возбуждения Н-холинорецепторов, преганглионарные синапсы имеют и М-холинорецепторы (активируются алкалоидом мускарином - мускариночувствительные), роль которых, повидимому, сводится к регуляции освобождения медиатора и чувствительности Н-холинорецепторов
Постганглионарные или периферические синапсы
text_fields
text_fields
arrow_upward
Постганглионарные или периферические синапсы, образуемые эфферентным проводником на эффекторе, отличаются у двух описываемых отделов вегетативной нервной системы.
2.1. Симпатические синапсы
Симпатические синапсы образуются не только в области многочисленных концевых ветвлений симпатического нерва, как у всех других нервных волокон, но и у мембран варикозов - многочисленных расширений периферических участков симпатических волокон в области иннервируемых тканей. Варикозы также содержат синаптические пузырьки с медиатором, хотя и в меньших концентрациях, чем терминальные окончания.
а) Медиатор симпатических синапсов — норадреналин
Основным медиатором симпатических синапсов является норадреналин и такие синапсы называют адренергическими. Рецепторы, связывающие адренергический медиатор получили название адренорецепторов. Различают два типа адренорецепторов - альфа и бета, каждый из которых делят на два подтипа - 1 и 2. Небольшая часть симпатических синапсов использует медиатор ацетилхолин и такие синапсы называют холинергическими, а рецепторы - холинорецепторами. Холинергические синапсы симпатической нервной системы обнаружены в потовых железах. В адренергических синапсах кроме норадреналина в существенно меньших количествах содержатся адреналин и дофамин, также относящиеся к катехоламинам, поэтому медиаторное вещество в виде смеси трех соединений раньше называли симпатином.
Синтез норадреналина из аминокислоты тирозина с помощью трех ферментов - тирозингидроксилазы, ДОФА-декарбоксилазы и дофамин-бета-гидроксилазы - происходит во всех частях постганглионарного нейрона: его теле, аксоне, варикозах и терминальных синаптических окончаниях. Однако из тела с током аксоплазмы поступает меньше 1% норадреналиш, основная же часть медиатора синтезируется в периферических отделах аксона и хранится и гранулах синаптических пузырьков. Содержащийся в гранулах норадреналин находится в двух фондах или пулах (запасных формах) - стабильном или резервном (85-90%) и лабильном, мобилизуемом в синаптическую щель при передаче возбуждения. Норадреналин лабильного фонда в случае необходимости медленно пополняется из стабильного пула. Пополнение запасов норадреналина, кроме процессов синтеза, осуществляется мощным обратным его захватом из синаптической щели пресинаптической мембраной (до 50% выделенного в синаптическую щель количества), после чего захваченный медиатор частично поступает в пузырьки, а не попавший в пузырьки - разрушается ферментом моноаминоксидазой (МАО).
б) Механизмы выделения норадреналина в синаптическую щель
Освобождение медиатора в синаптическую щель происходит квантами под влиянием импульса возбуждения, при этом число квантов пропорционально частоте нервных импульсов. Процесс высвобождения медиатора протекает с помощью экзоцитоза и является Са-зависимым.
Выделение норадреналина в синаптическую щель регулируется несколькими специальными механизмами:
1) связывание норадреналина в синаптической щели с альфа-2-адренорецепторами пресинаптической мембраны (рис.3.10), что играет роль отрицательной обратной связи и угнетает освобождение медиатора;
2) связывание норадреналина с пресинаптическими бета-адренорецепторами, что играет роль положительной обратной связи и усиливает освобождение медиатора.
При этом, если порции освобождающегося норадреналина небольшие, то медиатор взаимодействует с бета-адренорецепторами, что повышает его освобождение, а при высоких концентрациях медиатор связывается с альфа-2~адренорецептором, что подавляет его дальнейшее освобождение;
3) образование клетками эффектора и выделение в синаптическую щель простагландинов группы Е, подавляющих освобождение медиатора через пресинаптическую мембрану;
4) поступление в синаптическую щель адренергического синапса из рядом расположенного холинергического синапса ацетилхолина, связывающегося с М-холинорецептором пресинаптической мембраны и вызывающего подавление высвобождение норадреналина.
в) Судьба выделившегося в синаптическую щель норадреналина
Судьба выделившегося в синаптическую щель медиатора зависит от четырех процессов:
1) связывания с рецепторами пост- и пре-синаптических мембран,
2) обратного захвата пресинаптической мембраной,
3) разрушения в области рецепторов постсинаптической мембраны с помощью фермента катехол- О-метилтрансферазы (КОМТ),
4) диффузии из синаптической щели в кровоток, откуда норадреналин активно захватывается клетками многих тканей.
Диффундируя к постсинаптической мембране, норадреналин связывается с находящимися на ней адренорецепторами двух типов - альфа-1 и бета , образуя медиатор-рецепторный комплекс (рис.3.10).
Количество альфа-1 и бета-адренорецепторов в различных тканях неодинаково, например, в гладких мышцах артериальных сосудов внутренних органов преобладают альфа-адренорецепторы, а клетках миокарда - бета-адренорецепторы. Активация медиатором альфа- 1-адренорецепторов приводит к деполяризации и формированию ВПСП, более полого, низкоамплитудного и длительного, чем ВПСП нервных клеток и ПКП скелетных мышц. Стимуляция альфа-адренорецепторов вызывает также сдвиг метаболизма в мембране клеток и образование специфических молекул, называемых вторич ными посредниками медиаторного эффекта. Вторичными посредниками стимуляции альфа-адренорецепторов являются инозитол-3-фосфат и ионизированный кальций. Более подробно системы вторичных посредников будут рассмотрены в разделе, посвященном гуморальной регуляции функций.
Бета-адренорецепторы,
также как и альфа-, делят на 2 подтипа: бета-1
и бета-2.
Бета-1-адренорецепторы находятся в сердечной мышце и их стимуляция обеспечивает активацию основных физиологических свойств миокарда (автоматии, возбудимости, проводимости и сократимости).
Бета-2-адренорецепторы расположены в гладких мышцах артериальных сосудов, особенно скелетных мышц, коронарных артерий, бронхов, матки, мочевого пузыря и их стимуляция вызывает тормозной эффект в виде расслабления гладких мышц.
Хотя при этом и происходит гиперполяризация постсинаптической мембраны, выявить ТПСП не удается из-за очень медленного процесса и крайне низких амплитуд гиперполяризации. Стимуляция бета-адренорецепторов приводит в действие другую систему вторичных посредников - аденилатциклаза-цАМФ, причем считается, что бета-адренорецептор либо связан с аденилатциклазой, либо вообще является этим белком-ферментом.
Симпатическая нервная система является важнейшим регулятором обмена веществ в организме. С метаболическими эффектами симпатической нервной системы связано ее трофическое действие на ткани. Классическим экспериментальным подтверждением трофического влияния симпатической нервной системы является феномен Орбели — Гинецинского, суть которого состоит в следующем. Регистрируется амплитуда сокращений икроножной мышцы лягушки при раздражении иннервирующих ее передних корешков спинного мозга. Постепенно развивается утомление и амплитуда сокращений падает. Если в этот момент произвести раздражение симпатического пограничного ствола в этой области, то амплитуда сокращений восстанавливается, т.е. утомление исчезает
2.2. Парасимпатические синапсы
а) Медиатор симпатических синапсов – ацетилхолина
Парасимпатические постганглионарные или периферические синапсы используют в качестве медиатора ацетилхолин, который находится в аксоплазме и синаптических пузырьках пресинаптических терминалей в трех основных пулах или фондах. Это,
во-первых
, стабильный, прочно связанный с белком, не готовый к освобождению пул медиатора;
во-вторых
, мобилизационный, менее прочно связанный и пригодный к освобождению, пул;
в-третьих
, готовый к освобождению спонтанно или активно выделяемый пул. В пресинаптическом окончании постоянно происходит перемещение пулов с целью пополнения активного пула, причем этот процесс осуществляется и путем продвижения синаптических пузырьков к пресинаптической мембране, так как медиатор активного пула содержится в тех пузырьках, которые непосредственно прилежат к мембране. Освобождение медиатора происходит квантами, спонтанное выделение единичных квантов сменяется активным при поступлении импульсов возбуждения, деполяризующих пресинаптическую мембрану. Процесс освобождения квантов медиатора, также как и в других синапсах, является кальций-зависимым.
б) Механизм регуляции освобождения ацетилхолина в синаптическую щель
Регуляция освобождения ацетилхолина в синаптическую щель обеспечивается следующими механизмами:
1) Связыванием ацетилхолина с М-холинорецепторами пресинаптической мембраны, что оказывает тормозящее влияние на выход ацетилхолина - отрицательная обратная связь;
2) Связыванием ацетилхолина с Н-холинорецептором, что усиливает освобождение медиатора - положительная обратная связь;
3) Поступлением в синаптическую щель парасимпатического синапса норадреналина из рядом располагающегося симпатического синапса, что оказывает тормозной эффект на освобождение ацетилхолина;
4) Выделением в синаптическую шель под влиянием ацетилхолина из постсинаптической клетки большого числа молекул АТФ, которые связываются с пуринергическими рецепторами пресинаптической мембраны и подавляют освобождение медиатора - механизм, получивший название ретро-ингибирование.
(рис.3.11)
|
Рис.3.11. Парасимпатический синапс и его регуляция. |
1 — пресинаптическое окончание, 2 — синаптический пузырек, 3 — синаптическая щель с квантами ацетипхолина (АХ), 4 — постсинаптическая мембрана эффекторной клетки, 5 — рядом расположенный адренергический синапс. М — мускариновый холинорецептор, Н — никотиновый холинорецептор, ХЭ — холинэстераза, ГЦ-цГМФ — система вторичного посредника: гуанилатциклаза — циклический гуанозинмонофосфат, НА — норадреналин, (+) — стимуляция освобождения медиатора, (-) — подавление освобождения медиатора. |
в) Судьба выделившегося в синаптическую щель ацетилхолина
Выделившийся в синаптическую щель ацетилхолин удаляется из нее несколькими путями:
Во-первых
, часть медиатора связывается с холинорецепторами пост- и пресинаптической мембраны;
во-вторых
, медиатор разрушается ацетилхолинэстеразой с образованием холина и уксусной кислоты, которые подвергаются обратному захвату пресинаптической мембраной и вновь используются для синтеза ацетилхолина;
в-третьих
, медиатор путем диффузии выносится в межклеточное пространство и кровь, причем этот процесс происходит после связывания медиатора с рецептором. При удалении медиатора последним путем инактивируется почти половина выделившегося ацетилхолина.
На постсинаптической мембране ацетилхолин связывается с холинорецепторами, относящимися к М (мускариночувствительному) типу.
Образование на мембране медиатор-реиепторного комплекса приводит к общим для разных видов клеток реакциям:
во-первых
, к активации рецепторуправляемых ионных каналов и изменению заряда мембраны;
во-вторых
, к активации систем вторичных посредников в клетках.
В гладкомышечных и секреторных клетках желудочно-кишечного тракта, мочевого пузыря и мочеточника, бронхов, коронарных и легочных сосудов комплекс ацетилхолин-М-холино-рецептор активирует Na-каналы, приводит к деполяризации и формированию ВПСП, вследствие чего клетки возбуждаются и происходит сокращение гладких мышц или секреция пищеварительных соков. Этому же эффекту способствует активация вторичных посредников - инозитол-три-фосфата и ионизированного кальция. В то же время в клетках проводящей системы сердца, гладких мышцах сосудов половых органов комплекс ацетилхолин-М-холинорецептор активирует К-каналы и выходящий ток К+, приводя к гиперполяризации и тормозным эффектам - снижению автоматии, проводимости и возбудимости в миокарде, расширению артерий половых органов. Одновременно в этих клетках активируется система вторичных посредников - гуанилатциклаза-циклический гуанозинмонофосфат. Разнонаправленность эффектов парасимпатической регуляции при образовании на мембранах разных клеток комплекса ацетилхолин-М-холинорецептор дает основание предполагать наличие на постсинаптической мембране постганглионарных синапсов двух типов М-холинорецепторов, подобно типам адренорецепторов описанным выше. Вместе с тем, все М-холинорецепторы блокируются атропином, что снимает как парасимпатическую стимуляцию сокращения гладких мышц, так и парасимпатическое торможение деятельности сердца.
Эффективность синаптической передачи зависит от количества активных рецепторов на постсинаптической мембране, что отражает функции эффекторной клетки, синтезирующей мембранные рецепторы. Клетка эффектора регулирует число мембранных рецепторов в зависимости от интенсивности работы синапса, т.е. выделения в нем медиатора. Так, при перерезке вегетативного нерва (прекращении выделения медиатора) чувствительность иннервируемой им ткани к соответствующему медиатору возрастает из-за увеличения числа мембранных рецепторов, способных связывать-медиатор. Повышение чувствительности денервированных структур или сенситизация ткани является примером саморегуляции на уровне эффектора.
Взаимосвязи симпатической и парасимпатической регуляции функций
text_fields
text_fields
arrow_upward
Поскольку большинство эффектов симпатической и парасимпатической нервной регуляции являются противоположными, их взаимоотношения характеризуют иногда как антагонистические. Вместе с тем, существующие взаимосвязи между высшими вегетативными центрами и даже на уровне постганглионарных синапсов в тканях, получающих двойную иннервацию, позволяют применять понятие о реципрокной регуляции. Примером реципрокных взаимоотношений на уровне эффектора является акцентированный антагонизм или взаимоусиливающее противодействие.
Вегетативные ганглии играют значительную роль в распределении и распространении проходящих через них нервных импульсов. Число нервных клеток в ганглиях в несколько раз (в верхнем шейном спмпатическом узле в 32 раза, в ресничном узле в 2 раза) больше числа приходящих к ганглию преганглионарных волокон. Каждое из этих волокон образует синапсы на многих клетках ганглия. Поэтому нервные импульсы, поступающие по преганглионарному волокну в ганглий, могут оказывать влияние на большое число постганглионарных нейронов и, следовательно, на еще большее число мышечных и железистых клеток иннервируемого органа. Таким образом, достигается расширение зоны влияния преганглионарных волокон.
Насколько широко распространяются нервные импульсы, поступающие по преганглионарным волокнам, показывает тот факт, что у собак к нескольким крупным органам брюшной полости: к поджелудочной железе, тонким кишкам, надпочечникам и почкам подходит в среднем все около 70 волокон блуждающего нерва. Это ограниченное число волокон блуждающего нерва через свои синаптические связи с внутриорганными нервными узлами и сплетениями обеспечивает парасимпатическую иннервацию всех перечисленных внутренних органов.
На каждом постганглионарном нейроне имеются синапсы, образованные многими преганглионарными волокнами. С этим связано явление пространственной суммации нервных импульсов. Если раздражать одно преганглионарное волокно стимулами допороговой силы, то в постгаиглионарных волокнах не возникает потенциалов действия.
Если же раздражать несколько преганглионарных волокон стимулами той же силы, какая применялась для раздражения одного волокна, то обнаруживают потенциалы действия в постганглионарных волокнах в результате пространственной .
В симпатических нервных узлах зоны влияния отдельных преганглионарных волокон частично перекрывают друг друга. Поэтому при одновременном раздражении двух пучков этих волокон обнаруживается явление : эффект одновременного сверхпорогового раздражения двух пучков преганглионарных волокон всегда меньше, чем арифметическая сумма эффектов, возникающих при раздельном раздражении этих же пучков волокон.
При раздражении преганглионарных нервных волокон отчетливо обнаруживаются также явления временной суммации нервных импульсов. Раздражение одиночным стимулом, как правило, не дает эффекта даже при большой силе раздражения. (Неспособность возбуждаться под влиянием одиночных стимулов, свойственная нейронам вегетативной нервной системы и многим нейронам центральной нервной системы, называют итеративностью.) В отличие от этого в ответ на раздражение ритмическими стимулами преганглионарных волокон в постганглионарных нейронах возникает возбужденно вследствие временной суммации постсинаптических возбуждающих потенциалов.
Одностороннее проведение нервных импульсов в межнейронных синапсах, перекрытие зон влияния отдельных входящих в узел преганглионарных волокон, наличие временной и пространственной суммации и окклюзии показывают, что структурная организация и свойства нейронов и синапсов ганглиев вегетативной нервной системы таковы же, как нейронов и синапсов центральной нервной системы.
Характерной особенностью нейронов вегетативной нервной системы является относительно редкий ритм геперируемых ими импульсов, который не превышает 10-15 импульсов в секунду. Так, максимальный ритм импульсов, проходящих по сосудосуживающим нервным волокнам, не бывает чаще 6-8 в секунду. Частый ритм возбуждений преганглионарных волокон, превышающий частоту естественных импульсов, возникающих в нейронах вегетативной нервной системы, частично блокируется в синапсах, и постганглионарный нейрон возбуждается в более редком ритме. Таким образом, обнаруживается трансформация ритма нервных импульсов. Частота стимуляции свыше 100 в секунду вызывает полную блокаду проведения через синапс.
Являются ли ганглии вегетативной нервной системы рефлекторными центрами, т. е. происходит ли в них замыкание рефлекторных дуг? Этот вопрос многократно дебатировался, но до сих пор по является окончательно выясненным. В пользу такой возможности приводится тот факт, что в нервных сплетениях в некоторых органах, в частности в ауэрбаховом сплетении в желудке и в кишечнике, имеются обнаруженные Л. С. Догелем два типа нервных клеток, отличающихся по своей структуре и контактирующих между собой. Однако достаточно убедительных доказательств не приведено в пользу того, что в ганглиях и сплетениях вегетативной нервной системы имеются рецепторные нейроны, приносящие возбуждения от рецепторов и здесь же передающие импульсы к эффекторным нейронам.
Все функции организма условно делят на соматические и вегетативные. Первые связаны с деятельностью мышечной системы, вторые выполняются внутренними органами, кровеносными сосудами, кровью, железами внутренней секреции и т.д. Однако это деление условно, так как такая вегетативная функция, как обмен веществ, присуща скелетным мышцам. С другой стороны, двигательная активность сопровождается изменением функций внутренних органов, сосудов, желез.
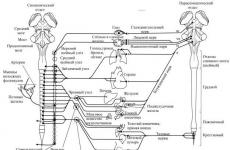
Вегетативной нервной системой называют совокупность нервных клеток спинного, головного мозга и вегетативных ганглиев, которые иннервируют внутренние органы и сосуды.
Вегетативная нервная система человека
Дуга вегетативного рефлекса отличается тем, что ее эфферентное звено имеет двухнейронное строение, т.е. от тела первого эфферентного нейрона, расположенного в ЦНС, идет преганглионарное волокно, которое заканчивается на нейронах вегетативного ганглия, расположенного вне ЦНС. От этого второго эфферентного нейрона идет постганглионарное волокно к исполнительному органу. Нервные импульсы по вегетативным рефлекторным дугам распространяются значительно медленнее, чем по соматическим. Во-первых, это обусловлено тем, что даже простейший вегетативный рефлекс является полисинаптическим, а большинство вегетативных нервных центров включает огромное количество нейронов и синапсов. Во-вторых, преганглионарные волокна относятся к группе «В», а постганглионарные – «С». Скорость проведения возбуждения по ним наименьшая. Все вегетативные нервы имеют значительно меньшую избирательность (например, n. Vagus), чем соматические.
Вегетативная нервная система делится на 2 отдела: симпатический и парасимпатический. Тела преганглионарных симпатических нейронов лежат в боковых рогах грудных и поясничных сегментов спинного мозга. Аксоны этих нейронов выходят в составе передних корешков и оканчиваются в паравертебральных ганглиях симпатических цепочек. От ганглиев идут постганглионарные волокна, иннервирующие гладкие мышцы органов и сосудов головы, грудной, брюшной полостей малого таза, а также пищеварительные железы. Существует симпатическая иннервация не только артерий и вен, но и артериол. В целом функция симпатической нервной системы состоит в мобилизации энергетических ресурсов организма за счет процессов диссимиляции, повышении его активности, в том числе и нервной системы.
Тела преганглионарных парасимпатических нейронов находятся в сакральном отделе спинного мозга, продолговатом и среднем мозге в области ядер III, VII, IX и X пар черепно-мозговых нервов. Идущие от них преганглионарные волокна заканчиваются на нейронах парасимпатических ганглиев. Они расположены около иннервируемых органов (параорганно) или в их толще (интрамурально). Поэтому постганглионарные волокна очень короткие. Парасимпатические нервы, начинающиеся от стволовых центров, также иннервируют органы и небольшое количество сосудов головы, шеи, а также сердце, легкие, гладкие мышцы и железы желудочно-кишечного тракта (ЖКТ). В ЦНС парасимпатических окончаний нет. Нервы, идущие от крестцовых сегментов, иннервируют тазовые органы и сосуды. Общей функцией парасимпатического отдела является обеспечение восстановительных процессов в органах и тканях, за счет усиления ассимиляции. Таким образом, сохраняется гомеостаз.
Высшие центры регуляции вегетативных функций находятся в гипоталамусе. Однако, на вегетативные центры влияет и кора больших полушарий. Это влияние определяется лимбической системой и центрами гипоталамуса. Многие внутренние органы имеют двойную, т.е. симпатическую и парасимпатическую иннервацию. Это сердце, органы ЖКТ, малого таза и другие. В этом случае влияние отделов вегетативной нервной системы носит антагонистический характер. Например, симпатические нервы усиливают работу сердца, тормозят моторику органов пищеварения, сокращают сфинктеры выводных протоков пищеварительных желез и расслабляют мочевой пузырь. Парасимпатические нервы влияют на функции этих органов противоположным образом. Поэтому в физиологических условиях функциональное состояние этих органов определяется преобладанием влияния того или иного отдела вегетативной нервной системы. Однако для организма их воздействие является синергичным. Например, такая функциональная синергия возникает при возбуждении барорецепторов сосудов, когда повышается артериальное давление. В результате их возбуждения повышается активность парасимпатических и снижается симпатических центров. Парасимпатические нервы уменьшают частоту и силу сердечных сокращений, а торможение симпатических центров приводит к расслаблению сосудов. Артериальное давление снижается до нормы. Во многих органах, имеющих двойную вегетативную иннервацию, постоянно преобладают регуляторные влияния парасимпатической нервной системы. Это железистые клетки ЖКТ, мочевой пузырь и другие. Есть органы, имеющие только одну иннервацию. Например, большинство сосудов иннервируется только симпатическими нервами, которые постоянно поддерживают их в суженном состоянии, т.е. тонусе.
В 80-х годах А.Д. Ноздрачевым сформулирована концепция метасимпатической нервной системы. Согласно ей, интрамуральные ганглии вегетативной нервной системы, образующие нервные сплетения, являются простыми нейронными сетями, аналогичными ядрам ЦНС. В этих небольших нейронных скоплениях, преимущественно находящихся в стенке органов пищеварительного канала, происходит восприятие раздражения, переработка информации и передача к эфферентным нейронам, а затем исполнительным органам. Ими являются гладкомышечные клетки пищеварительного канала, матки, кардиомиоциты., т.е. ганглии достаточно автономны от ЦНС. Однако сигналы от них поступают и в ЦНС, перерабатываются в ней, а затем через экстрамуральные парасимпатические нервы передаются на эфферентные нейроны ганглия, а от него на исполнительный орган, т.е. эфферентные нейроны ганглиев являются общим конечным путем и для экстрамуральных парасимпатических нервов и для других нейронов ганглиев.
В стенке пищевода, желудка, кишечника имеется 3 связанных между собой сплетения: подсерозное, межмышечное (ауэрбахово), подслизистое (мейснерово). Клетки, составляющие сплетения относятся по классификации А.С. Догеля к трем типам:
I тип – нейроны с многочисленными короткими дендритами и длинными аксонами. Аксон заканчивается на гладкомышечных клетках и железистых клетках пищеварительного канала. Эти нейроны являются эффекторными.
II тип – более крупные нейроны, имеющие несколько дендритов и короткий аксон, образующий синапс на нейронах первого типа. Окончания дендритов находится в подслизистой и слизистой оболочках, т.е. эти клетки являются чувствительными.
III тип – служат для передачи сигналов между другими нейронами ганглиев. Их можно считать ассоциативными, т.е. интернейронами. Их меньше других.
Кроме того, в сплетениях выделяют так называемые нейроны-генераторы. Они обладают автоматией и задают частоту ритмической активности гладких мышц ЖКТ.
Таким образом, отличительной особенностью метасимпатической нервной системы является то, что ее эфферентные нейроны всегда расположены интрамурально и регулируют частоту ритмических сокращений сердца, кишечника, матки и т.д. Поэтому, даже после перерезки всех экстрамуральных нервов, идущих к этим органам, их нормальная функция сохраняется.
Наличие метасимпатической системы способствует освобождению ЦНС от излишней информации, так как метасимпатические рефлексы замыкаются в интрамуральных ганглиях. Она обеспечивает поддержание гомеостаза, управляя работой тех внутренних органов, которые имеют ее.
Регуляция функций вегетативной нервной системой осуществляется по рефлекторному принципу, т.е. раздражение периферических рецепторов приводит к возникновению нервных импульсов, которые после анализа и синтеза в вегетативных центрах поступают на эфферентные нейроны, а затем исполнительные органы. Поэтому все вегетативные рефлексы, в зависимости от участка рецепторного и эфферентного звена, делятся на следующие группы:
1. Висцеро-висцеральный. Это рефлексы, которые возникают вследствие раздражения интерорецептолров внутренних органов и проявляются изменениями их функций. Например, при механическом раздражении брюшины или органов брюшной полости происходит урежение и ослабление сердечных сокращений (рефлекс Гольца).
2. Висцеро-дермальные. Раздражение интерорецепторов внутренних органов, приводит к изменению потоотделения, просвета сосудов кожи, кожной чувствительности.
3. Сомато-висцеральные . Действие раздражителя на соматические рецепторы, например, рецепторы кожи, приводит к изменению деятельности внутренних органов. К этой группе относятся рефлексы Данини-Ашнера (урежение сердцебиений при надавливании на глазные яблоки).
4. Висцеро-соматические . Раздражение интерорецепторов вызывает изменение двигательных функций. Возбуждение хеморецепторов сосудов углекислым газом, способствует усилению сокращений межреберных дыхательных мышц. Пари нарушении механизмов вегетативной регуляции возникают изменения висцеральных функций. В частности, психосоматические заболевания.